Lymph nodes are highly organized organs and essential for regulation of the adaptive immune system. Although lymph nodes are highly organized, embryonic lymphoid node formation starts with the interaction of a subset of type 3 Innate Lymphoid Cells (ILC3), previously named Lymphoid Tissue Inducer (LTi) cells, with mesenchymal organizer cells at pre-determined locations throughout the body (Figure 1).
The ILC family has been recently described and is now recognized as essential for several immunological functions. Interestingly, these innate cells are essential for development of the adaptive immune system by their involvement in the embryonic formation of lymph nodes.
Previously, I have shown that retinoic acid is essential for the differentiation of both the ILC3 as the mesenchymal organizer cells and hence essential for embryonic lymph node formation. More importantly, altering levels of the raw material for retinoic acid, vitamin A, within the diet of the pregnant mouse affected the size of lymph nodes in her offspring at adult age. Therefore, the diet of the pregnant mother is important for the proper development of the immune system within the embryos and has a lifelong effect on the immunity of her offspring. Basically, you are what your mother ate.
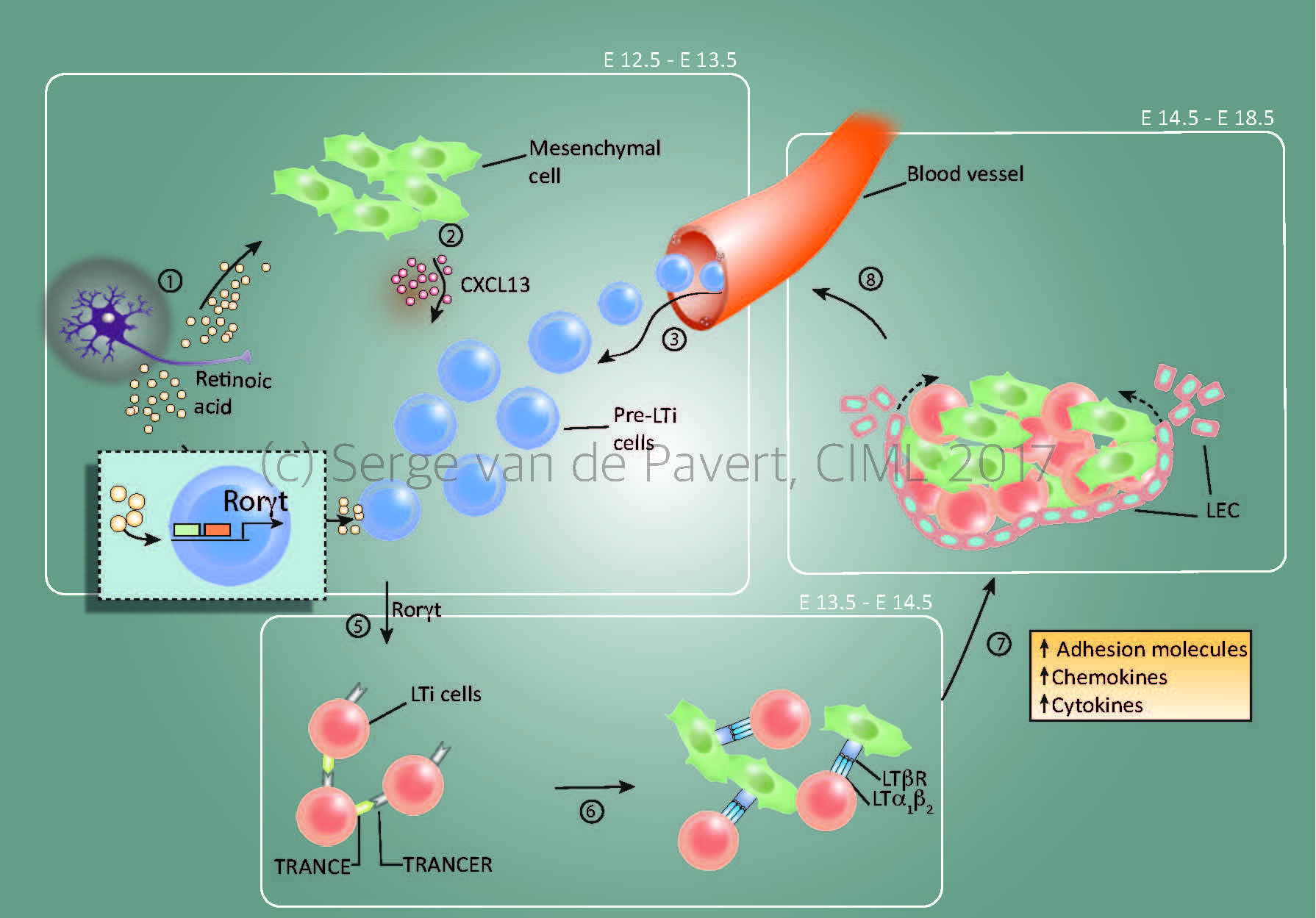
Figure 1: Lymph node formation within the mouse embryo starts around E12.5 by Cxcl13 secretion by resident mesenchymal cells, under influence by retinoic acid. The stage at which the lymph node anlagen is initiated depends on the location within the embryo, with the most anterior lymph nodes (E12.5= cervical, mandibular, axillary) forming the first and the most posterior as last (E14.5/15.5= inguinal, popliteal). The presence of Cxcl13 and possibly other chemokines expressed by the mesenchymal cells, attract (pre-) Lymphoid Tissue Inducer (LTi) cells towards the niche. Within this niche, retinoic acid induces the essential nuclear transcription factor RORγt within the pre-LTi cells so that they mature into LTi cells, which can interact with the mesenchymal/stromal organiser cells via the lymphotoxin signalling pathway.
Expansion of the lymph node anlagen takes place after lymphotoxin signalling started (around E14.5 for the most anterior lymph nodes), with the attraction of more LTi cells, and thus more interactions with resident mesenchymal cells. Also, the mesenchymal cells produce Vegfc, a potent attraction molecule for lymphatic endothelial cells, which form a layer of lymphatic endothelial cells around the lymph node anlagen and eventually connect it to the lymphatic vasculature. At this stage, the chemokine Ccl21 expressed by the lymphatic endothelial cells and vasculature are important for the transport of LTi cells towards the lymph node anlagen.
It is still unclear how lymph nodes are formed at specific locations, and possibly the nerve fibers and blood vessel bifurcations could play a role in establishing their location. Our aim is study the anatomical correlation of the structures in relation to lymph nodes and their associated lymphatic vasculature, and to study the role of nerve fibers in this process. This will be done by the use of light-sheet microscopy, which enables the whole mount acquisition of fluorescently stained mouse embryos up to E13.5 (Figure 2).
Also, my group will study other substances in the maternal diet that affect differentiation of key players involved in lymph node formation. These studies will provide insights into the ontogeny and functioning of the ILC3 family and provide knowledge on the requirements for the formation of embryonic lymph nodes.
The overall objective of my group is to characterize all players involved in development of the immune system, specifically in relation to the formation of lymph nodes and lymphatic vasculature.
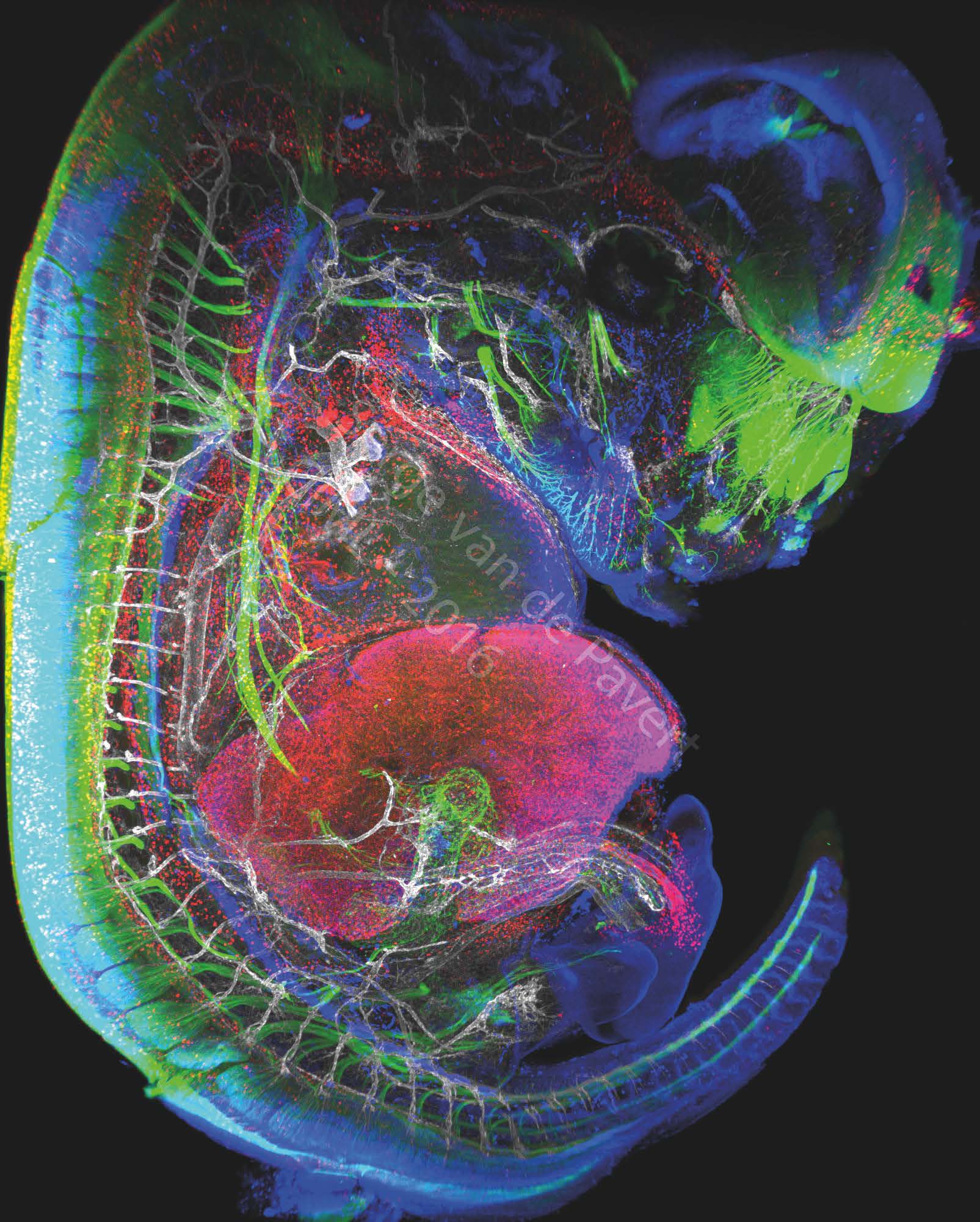
Figure 2: Immunofluorescent staining of a complete mouse E13.5 embryo reveals neurons (labeled with BetaIII Tubulin), macrophages and lymphatic endothelial cells in red (labeled with Lyve-1), the nuclei of specific neurons and lymphatic endothelial cells are marked in blue (Prox1 labeling) and the vascular network in white (Pecam-1 labeling). The eye of the embryo and the nasal neurons can be seen in green. On the left, the spinal cord is marked in green. The vertebrae are visible through the marking of the dorsal root ganglia in green, and blood vessels marked in white. The heart is visible just above the liver which is labeled with LYVE-1 antibody. The embryonic intestine, not far from the liver, is visualized by the innervation of neurons in green. The umbilical cord can be observed by neighboring white blood vessels and macrophages in red. This picture was generated in the study on the formation of lymph nodes and lymphatic system. Their formation, initiated in the embryonic stage, can be modified by changing certain dietary factors affecting cell differentiation. These visualizations are therefore intended to analyze the movements of cells involved in the formation of the lymphatic system.
Image: Serge van de Pavert/CIML